Физиология нервов и нервных волокон
Физиология нервов и нервных волокон
Нервные волокна выполняют специализированную функцию – проведение нервных импульсов. По морфологическому признаку волокна делятся на миелиновые (покрытые миелиновой оболочкой) и безмиелиновые. Нерв состоит из большого числа нервных волокон (миелиновых и безмиелиновых), заключенных в общую оболочку.
Нервное волокно обладает следующими свойствами: возбудимостью, проводимостью и лабильностью.
Распространение возбуждения по нервным волокнам осуществляется на основе ионных механизмов генерации потенциала действия. При распространении возбуждения по безмиелиновому нервному волокну местные электрические токи, которые возникают между его возбужденным участком, заряженным отрицательно, и невозбужденными, Заряженными положительно, деполяризуют мембрану до критического уровня, что приводит к генерации ПД в соседних невозбужденных участках, которые становятся возбужденными, и т. д. Этот процесс происходит в каждой точке мембраны на всем протяжении волокна. Такое проведение возбуждения называется непрерывным. Возбуждение по нервному волокну может распространяться в обе стороны от места его возникновения. Если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение, то возбуждение зафиксируют электроды по обе стороны от места раздражения.
Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки (перехватов Ранвье), приводит к тому, что местные электрические токи не могут проходить через миелин, они возникают только между соседними перехватами Ранвье, где деполяризуют мембрану невозбужденного перехвата и генерируют ПД (рис.4). Возбуждение как бы «перепрыгивает» через участки нервного волокна, покрытые миелином. Такой механизм распространения возбуждения называется сальтаторным, или скачкообразным, он позволяет более быстро и экономично передавать информацию по сравнению с непрерывным проведением, поскольку в него вовлекается не вся мембрана, а только ее небольшие участки.
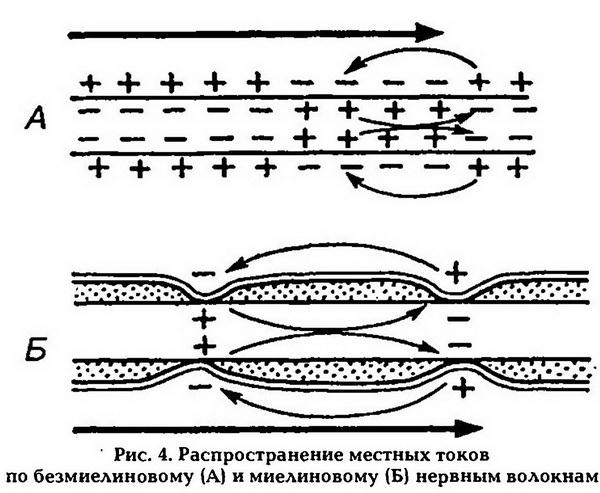
Амплитуда ПД в 5 6 раз превышает пороговую величину, не-обходимую для возбуждения соседнего перехвата, поэтому ПД может «перепрыгивать» не только через один, но и через несколько перехватов. Это явление может наблюдаться при снижении возбудимости соседнего перехвата под действием какого-либо фармакологического вещества, например, новокаина, кокаина и др.
Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность. Различные факторы, изменяющие свойства волокон (наркотические вещества, охлаждение, перевязка и т. д.), приводят к нарушению передачи возбуждения.
Возбуждение по нервному волокну, входящему в состав нерва, распространяется изолированно, т. е. не переходя с одного волокна на другое. Это обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон, и основная часть тока, возникающего между возбужденным и невозбужденным участками, проходит по межклеточной жидкости, не действуя на другие волокна. Если бы возбуждение передавалось с одного нервного волокна на другое, то нормальное функционирование организма было бы невозможно, так как нервы содержат большое количество чувствительных, двигательных, вегетативных волокон, которые несут информацию как от различных рецепторов к ЦНС, так и от ЦНС к эффекторным органам.
Нервные волокна по скорости проведения возбуждения делятся на три типа: А, В, С. Волокна типа А, в свою очередь, делятся на подтипы: А-а, А-р, А-у, А-5 (рис. 5).
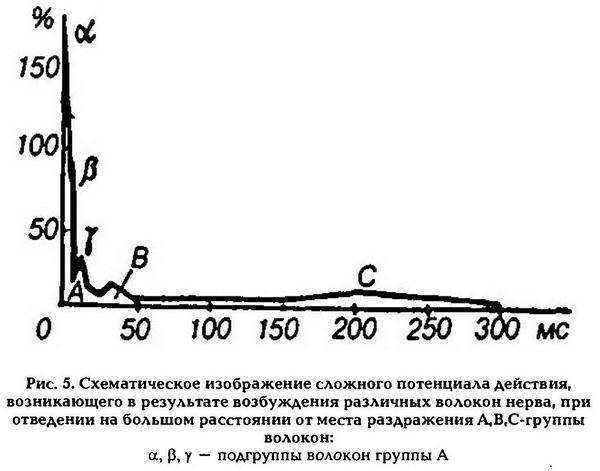
Волокна типа А покрыты миелиновой оболочкой. Наиболее толстые из них А-а имеют диаметр 12 – 22 мкм и скорость проведения возбуждения 70–120 м/с. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от рецепторов мышц к соответствующим нервным центрам.
Три другие группы волокон типа А (р, у, 5) имеют меньший диаметр – от 8 до 1 мкм и меньшую скорость проведения возбуждения – от 5 до 70 м/с. Волокна этих групп преимущественно проводят возбуждение от различных рецепторов (тактильных, температурных, болевых, рецепторов внутренних органов) в ЦНС, за исключением у-волокон, значительная часть которых проводит возбуждение от спинного мозга к интрафузальным мышечным волокнам.
К волокнам типа В относятся миелинизироваыные преганглионарные волокна вегетативной нервной системы. Их диаметр – 1 – 3,5 мкм, а скорость проведения возбуждения – 3–18 м/с.
К волокнам типа С относятся безмиелиповые нервные волокна малого диаметра – 0,5 – 2 мкм. Скорость проведения возбуждения в этих волокнах не более 3 м/с (0,5 – 3 м/с). Большинство волокон типа С – это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
Нервные волокна обладают лабильностью (функциональной подвижностью) – способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в соответствии с ритмом раздражения без искажений. Лабильность определяется длительностью потенциала действия (длительностью фазы абсолютной рефрактерности), у нервных волокон лабильность очень высокая (до 1000 Гц).
Н. Е. Введенский (1891 г.) обнаружил, что если участок нерва подвергнуть воздействию повреждающего агента (химического вещества, нагревания или охлаждения, постоянного тока), то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не может воспроизвести ритм раздражения – проведение импульсов нарушается. Такое состояние было названо парабиозом. В развитии парабиоза различают три последовательно сменяющие друг друга фазы: уравнительную, парадоксальную, тормозную.
В уравнительную фазу ответные реакции на частые и редкие раздражители становятся одинаковыми. В нормальных условиях величина ответной реакции иннервируемых нервом мышечных волокон зависит от частоты раздражения: на редкие раздражители ответная реакция меньше, а на частые – больше. В начальную стадию парабиоза при редком ритме раздражений (25 Гц) все импульсы проводятся через поврежденный участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражений (100 Гц) последующие импульсы могут поступать в период рефрактерности, поэтому часть импульсов не проводится. Например, если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100), то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) – происходит уравнивание ответной реакции.
В парадоксальную фазу происходит дальнейшее снижение лабильности. Ответная реакция возникает и на редкие, и на частые раздражители, но на частые она меньше, так как они еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. В результате ответная реакция на редкие раздражители будет больше, чем на частые.
В тормозную фазу и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна де-поляризована и не способна генерировать ПД, т. е. нерв утрачивает способность к проведению возбуждений.
Явление парабиоза лежит в основе локального обезболивания. Влияние анестезирующих веществ связано с нарушением механизма проведения возбуждения по нервным волокнам и снижением лабильности. Парабиоз – явление обратимое. Если парабнотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, по в обратной последовательности.
Возникновение парабиотического состояния связано с тем, что при действии на нервное волокно парабиотического фактора нарушается способность мембраны увеличивать натриевую проницаемость (инактивация натриевых каналов) в ответ на раздражение, и проведение следующего импульса блокируемся.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Очищение кишечника с помощью растительных волокон
Очищение кишечника с помощью растительных волокон Волоконца хорошо перемолотого, а затем пропаренного шиповника или гранулы морской травы ламинарии (в сухом виде) принимать лучше всего на ночь, спустя 2—3 часа после ужина, по 1/2 ч. ложки. Проходя через кишечник, они
ЛЕКЦИЯ № 15. Средства, действующие в области чувствительных нервных окончаний. Средства, понижающие чувствительность нервных окончаний
ЛЕКЦИЯ № 15. Средства, действующие в области чувствительных нервных окончаний. Средства, понижающие чувствительность нервных окончаний 1. Местноанестезирующие средства Препараты этой группы избирательно блокируют процесс передачи возбуждения в эфферентных нервах и
ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон 1. Физиология нервов и нервных волокон. Типы нервных волокон Физиологические свойства нервных волокон:1) возбудимость – способность приходить в состояние возбуждения в ответ на раздражение;2) проводимость
6. Физиология нервов и нервных волокон. Типы нервных волокон
6. Физиология нервов и нервных волокон. Типы нервных волокон Физиологические свойства нервных волокон:1) возбудимость – способность приходить в состояние возбуждения в ответ на раздражение;2) проводимость – способность передавать нервные возбуждение в виде потенциала
Поражения нервных корешков
Поражения нервных корешков Симптоматика корешкового синдрома зависит от расположения поражения корешков: шейно-грудной радикулит, вертебро-пояснично-крестцовый радикулит.При шейно-грудном радикулите появляются боли различной интенсивности, которые иррадиируют из
Физиология нервов и нервных волокон
Физиология нервов и нервных волокон Нервные волокна выполняют специализированную функцию – проведение нервных импульсов. По морфологическому признаку волокна делятся на миелиновые (покрытые миелиновой оболочкой) и безмиелиновые. Нерв состоит из большого числа
Свойства нервных процессов
Свойства нервных процессов Под свойствами нервных процессов понимают такие характеристики возбуждения и торможения, как сила, уравновешенность и подвижность этих процессов.Сила нервных процессов. При измерении силы процесса возбуждения обычно пользуются кривой
13. Какова роль пищевых волокон при запорах и зачем есть отруби?
13. Какова роль пищевых волокон при запорах и зачем есть отруби? Прежде всего, определим, что такое пищевые волокна. Пищевыми волокнами называют ту часть растительных компонентов пищи, которая устойчива к действию пищеварительных соков, т. е. они не перевариваются в
ПРОФИЛАКТИКА И ЛЕЧЕНИЕ НЕРВНЫХ ЗАБОЛЕВАНИЙ
ПРОФИЛАКТИКА И ЛЕЧЕНИЕ НЕРВНЫХ ЗАБОЛЕВАНИЙ Нормализация энергетического соотношения инь и ян в меридиане
ВОДОЛЕЧЕНИЕ ПРИ НЕРВНЫХ ЗАБОЛЕВАНИЯХ
ВОДОЛЕЧЕНИЕ ПРИ НЕРВНЫХ ЗАБОЛЕВАНИЯХ Я уже говорила о том, что расстроенные нервы прекрасно излечиваются при помощи релаксации, наблюдения за морем, полного расслабления. Но можно излечить подобные заболевания через применение морской воды.В наше время стрессов и
Значение растительных волокон
Значение растительных волокон Одно из основных преимуществ сложных углеводов — волокна. В стенках растительных клеток находятся разные виды волокон. Они играют важнейшую роль в процессе пищеварения, уменьшают вероятность того, что различные канцерогены и иные опасные
Жизнь в эпоху нервных перегрузок
Жизнь в эпоху нервных перегрузок Доктор Бейтс подчеркивал: «Чтобы убедиться в том, что напряжение ухудшает зрение, подумайте о чем-нибудь неприятном, о каком-нибудь чувстве физического дискомфорта или о чем-нибудь плохо видимом. Когда глаза откроются, будет обнаружено,
Заговор от нервных припадков
Заговор от нервных припадков Из письма:«Дорогая Наталья Ивановна, помогите мне, пожалуйста, если сможете. Ваши книги мне в руки попались случайно, я почитала письма людей и Ваши ответы и решила написать Вам. Вы показались мне человеком мудрым и добрым, прошу Вас, не
Майоран при нервных расстройствах, депрессии, бессоннице
Майоран при нервных расстройствах, депрессии, бессоннице Потребуется:Майоран (трава) – 1 ст. л.Кипяток – 1 стаканЧто делать:Майоран залить кипятком. Настоять 30 минут. Процедить. Эта порция на 1 день (выпить в три
Увеличение проводимости мышечных волокон
Увеличение проводимости мышечных волокон Нервная система управляет скелетными мышцами через сеть нейронов, которые связаны с мышечными волокнами через специальные соединения. Нервный импульс (командный сигнал) может активизировать все или некоторые из волокон с
Укрепление мышечных волокон
Укрепление мышечных волокон Как мы уже говорили, в мышцах может происходить преобразование одного типа волокон в другое. Считается, что короткие, интенсивные регулярные упражнения увеличивают скорость преобразования волокон медленного сокращения в волокна быстрого