Биоэлектрические явления в возбудимых тканях. Природа возбуждения
Биоэлектрические явления в возбудимых тканях. Природа возбуждения
Возбуждение представляет собой сложную совокупность физических, химических и физико-химических процессов, в результате которых происходит быстрое и кратковременное изменение электрического потенциала мембраны.
Первые исследования электрической активности живых тканей были проведены Л. Гальвани. Он обратил внимание на ссн сращение мышц препарата задних лапок лягушки, подвешенной на медном крючке, при соприкосновении с железными перилами балкона (первый опыт Гальвани). На основании этих наблюдений им был сделал вывод, что сокращение лапок вызвано «животным электричеством», которое возникает в спинном мозге и передается по металлическим проводникам (крючку и перилам) к мышцам.
Физик А. Вольта, повторив этот опыт, пришел к другому заключению. Источником тока, по его мнению, является не спинной мозг и «животное электричество», а разность потенциалов, образующаяся в месте контакта разнородных металлов – меди и железа, а нервно-мышечный препарат лягушки является лишь проводником электричества. В ответ на эти возражения Л. Гальвани усовершенствовал опыт, исключив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки лягушки, затем набрасывал нерв на мышцы голени, что вызывало сокращение мышцы (второй опыт Гальвани), тем самым доказав существование «животного электричества».
Позднее Дюбуа-Реймоном было установлено, что поврежденный участок мышцы имеет отрицательный заряд, а неповрежденный участок – положительный. При набрасывании нерва между поврежденным и неповрежденным участками мышцы возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током покоя, или током повреждения. Так было показано, что наружная поверхность мышечных клеток заряжена положительно по отношению к внутреннему содержимому.
Мембранный потенциал
В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая называется мембранным потенциалом [МП), или, если это клетка возбудимой ткани, – потенциалом покоя. Так как внутренняя сторона мембраны заряжена отрицательно по отношению к наружной, то, принимая потенциал наружного раствора за нуль, МП записывают со знаком «минус». Его величина у разных клеток колеблется от минус 30 до минус 100 мВ.
Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном (1902). Исходя из того, что мембрана клеток обладает высокой проницаемостью для ионов калия и малой проницаемостью для других ионов, он показал, что величину мембранного потенциала можно определить, используя формулу Нернста.
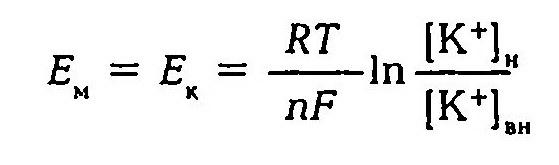
В 1949–1952 гг. А. Ходжкин, Э. Хаксли, Б. Катц создали со-временную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30 -50 раз больше ионов калия, в 8–10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Проницаемость мембраны для ионов обусловлена ионными каналами, макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, другие (потенциалозависимые) открываются и закрываются в ответ на изменения МП. Потенциалозависимые каналы подразделяются на натриевые, калиевые, кальциевые и хлорные. В состоянии физиологического покоя мембрана нервных клеток в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Таким образом, согласно обновленной мембранной теории асимметричное распределение ионов по обе стороны мембраны и связанное с этим создание и поддержание мембранного потенциала обусловлено как избирательной проницаемостью мембраны для различных ионов, так и их концентрацией по обе стороны от мембраны, а более точно величину мембранного потенциала можно рассчитать по формуле.
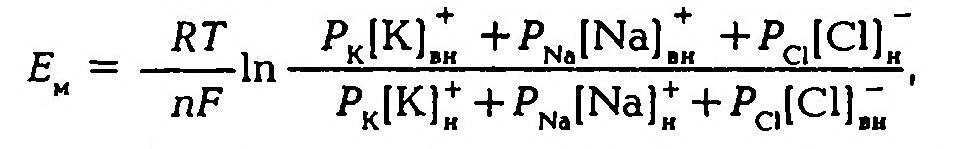
Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т. е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы – крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд. Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки. То что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей 1. Физиологическая характеристика возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и
1. Физиологическая характеристика возбудимых тканей
1. Физиологическая характеристика возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей.Раздражители – это
2. Законы раздражения возбудимых тканей
2. Законы раздражения возбудимых тканей Законы устанавливают зависимость ответной реакции ткани от параметров раздражителя. Эта зависимость характерна для высоко организованных тканей. Существуют три закона раздражения возбудимых тканей:1) закон силы
3. Понятие о состоянии покоя и активности возбудимых тканей
3. Понятие о состоянии покоя и активности возбудимых тканей О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма, нет
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.Процессы метаболизма в
2. Основные характеристики и законы возбудимых тканей
2. Основные характеристики и законы возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей.Раздражители – это
3. Понятие о состоянии покоя О и активности возбудимых тканей
3. Понятие о состоянии покоя О и активности возбудимых тканей О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма.Основные
2 глава Патологические изменения в тканях
2 глава Патологические изменения в тканях При заболеваниях различных органов и систем в коже, соединительной ткани, миофасциальных структурах, надкостнице образуются патологические изменения , которые выявляются при наружном осмотре, поверхностной и глубокой
2 глава Патологические изменения в тканях
2 глава Патологические изменения в тканях При заболеваниях различных органов и систем в коже, соединительной ткани, миофасциальных структурах, надкостнице образуются патологические изменения, которые выявляются при наружном осмотре, поверхностной и глубокой
Глава 2. Физиология возбудимых тканей
Глава 2. Физиология возбудимых тканей Способность адаптироваться к постоянно изменяющимся условиям внешней среды является одним из основных признаков живых систем. В основе приспособительных реакций организма лежит раздражимость – способность реагировать на
Законы раздражения возбудимых тканей
Законы раздражения возбудимых тканей Эти законы отражают определенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани. К законам раздражения относятся: закон силы, закон «все или ничего», закон раздражения Дюбуа-Реймона (аккомодации),
Газообмен в легких и тканях
Газообмен в легких и тканях В легких происходит газообмен между поступающим в альвеолы воздухом и протекающей по капиллярам кровью. Интенсивному газообмену между воздухом альвеол и кровью способствует малая толщина описанного аэрогематического барьера. Альвеолярный
Инородное тело в тканях
Инородное тело в тканях Рыбья кость, иголки, осколки костей —
Абсцесс (скопление гноя в тканях)
Абсцесс (скопление гноя в тканях) • Взять по 20 г порошка клубней топинамбура, семян льна и травы донника желтого, по 10 г листьев мальвы, топинамбура и алтея; 2 ст. л. смеси залить 300 г кремниевой воды, варить 5 мин, настоять, укутав, 1 час, процедить, добавить 1 ст. л. меда.
Абсцесс, скопление гноя в тканях
Абсцесс, скопление гноя в тканях ? Взять по 20 г семян льна и травы донника, по 10 г листьев мальвы и алтея; 2 ст. л. смеси залить 300 мл воды, всыпать 1 ст. л. пищевой соды, варить 5 мин. Для компрессов.? Взять по 1 ст. л. луковиц лилии чисто-белой, корневищ купены душистой и корневищ
Абсцесс, скопление гноя в тканях
Абсцесс, скопление гноя в тканях — Взять по 20 г семян льна и травы донника лекарственного, по 10 г листьев мальвы и алтея; 2 ст. ложки смеси залить 300 г воды, варить 5 мин., охладить, процедить, добавить 3 ст. ложки яблочного уксуса. Для