Физиология мышц
Физиология мышц
Существует три типа мышц: поперечно-полосатые скелетные мышцы, поперечно-полосатая сердечная мышца и гладкие мышцы.
Мышцы обладают следующими физиологическими свойствами:
1. возбудимостью, т. е. способностью возбуждаться при действии раздражителей;
2. проводимостью – способностью проводить возбуждение;
3. сократимостью – способностью изменять свою длину или напряжение при возбуждении;
4. растяжимостью – способностью изменять свою длину под действием растягивающей силы;
5. эластичностью – способностью восстанавливать свою первоначальную длину после прекращения растяжения.
Сила мышцы определяется максимальным грузом, который мышца может поднять. Мышцы способны совершать работу. Работа мышц определяется произведением величины поднятого груза на высоту подъема. Максимальная работа производится при средних величинах нагрузок. Лабильность мышцы равна 200–300 Гц.
При непосредственном раздражении мышцы (прямое раздражение) или опосредованно через иннервирующий ее двигательный нерв (непрямое раздражение) одиночным стимулом возникает одиночное мышечное сокращение, в котором выделяют три фазы: латентный период. – время от начала действия раздражителя до начала ответной реакции; фазу сокращения (фаза укорочения) и фазу расслабления.
В естественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, на которые мышца отвечает длительным сокращением. Длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение, называется тетаническим сокращением, или тетанусом (рис.6). Различают два вида тетануса: зубчатый и гладкий.

Если каждый последующий стимул поступает к мышце в тот период, когда она находится в фазе укорочения, то возникает гладкий тетанус, а если в фазу расслабления – зубчатый тетанус.
Амплитуда тетанического сокращения превышает амплитуду одиночного мышечного сокращения. Н. Е. Введенский объяснил это явление фазными изменениями возбудимости мышцы, введя понятие об оптимуме и пессимуме частоты раздражения. Оптимум – такая частота раздражения, при которой каждое последующее раздражение наносится в фазу повышенной возбудимости. Тетанус при этом будет максимальным по амплитуде. Пессимум – такая частота раздражения, при которой каждое последующее раздражение наносится в фазу пониженной возбудимости. Амплитуда тетануса при этом будет минимальной.
Различают несколько видов мышечных сокращений: изотонический, изометрический и смешанный. При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, если отсутствует сопротивление изменению ее длины. К изотоническому типу сокращений относятся сокращения мышц языка. При изометрическом сокращении длина мышечных волокон остается постоянной, а их напряжение возрастает. Такое сокращение мышцы возникает при попытке поднять чрезмерно большой груз. В естественных условиях сокращения мышц никогда не бывают чисто изотоническими или изометрическими, они имеют смешанный характер, т. е. происходит изменение и длины, и напряжения мышцы.
Механизм мышечного сокращения
Мышцы состоят из мышечных волокон, а те – из множества тонких нитей – миофибрилл, расположенных продольно. Каждая миофибрилла состоит из нитей сократительных белков актина и миозина. Перегородки, называемые Z-пластинами, разделяют миофибриллы на участки – саркомеры. В саркомере чередуются поперечные светлые и темные полосы. Поперечная исчерченность миофибрилл обусловлена определенным расположением нитей актина и миозина. В центральной части каждого саркомера расположены толстые нити миозина. На обоих концах саркомера находятся тонкие нити актина, прикрепленные к Z-пластинам. Нити миозина выглядят в световом микроскопе как светлая полоска (Н-зона) в темном диске, который содержит нити миозина и актина и называется анизотропным, или А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и кажутся светлыми, они называются изотропными, или I-дисками. По их середине проходит темная линия – Z-мембрана. Благодаря такому периодическому чередованию светлых и темных дисков сердечная и скелетная мышцы выглядят поперечно-полосатыми (рис.7).
В состоянии покоя концы толстых и тонких нитей лишь незначительно перекрываются на уровне А-диска. При сокращении тонкие актиновые нити скользят вдоль толстых миозиновых нитей, двигаясь между ними к середине саркомера. Сами актиновые и миозиновые нити своей длины не изменяют. Миозиновые нити имеют поперечные мостики (выступы) с головками, которые отходят от нити биполярно. Актиновая нить состоит из двух закрученных одна вокруг другой цепочек молекул актина. На нитях актина расположены молекулы тропонина, а в желобках между двумя актина лежат нити тропомиозина. Молекулы тропомиозина в покое располагаются так, что предотвращают прикрепление поперечных мостиков миозина к актиновым нитям.
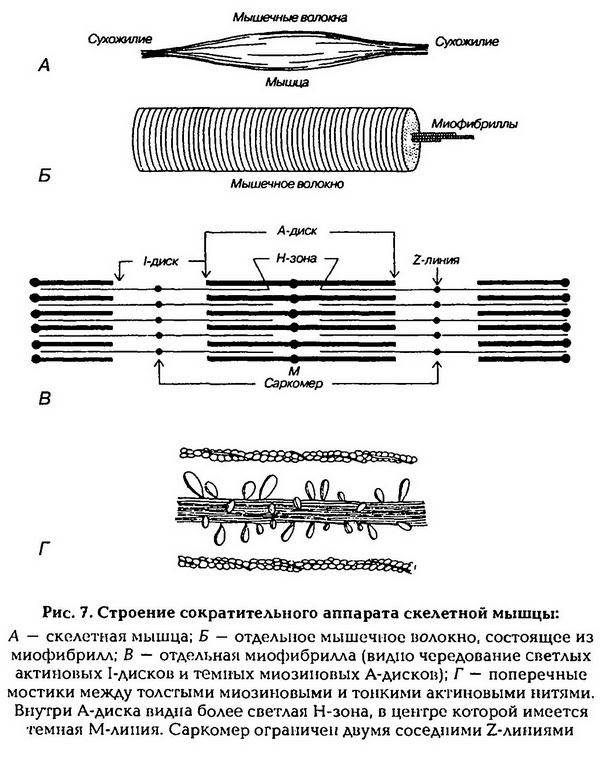
Во многих местах участки поверхностной мембраны мышечной клетки углубляются в виде трубочек внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-систему). Параллельно миофибриллам и перпендикулярно поперечным трубочкам расположена система продольных трубочек (альфа-система). Пузырьки на концах этих трубочек, в которых сосредоточено основное количество внутриклеточного кальция, подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В состоянии покоя миозиновый мостик заряжен энергией (миозин фосфорилирован), но он не может соединиться с нитью актина, так как между ними находится система из нитей тропомиозина и молекул тропонина. При возбуждении ПД распространяется по мембранам Тсистемы внутрь клетки и вызывает высвобождение ионов кальция из альфа-системы. С появлением ионов кальция в присутствии АТФ происходит изменение пространственного положения тропонина – нить тропомиозина сдвигается и открываются участки актина, присоединяющие миозиновые головки. Соединение головки фосфорилированного миозина с актином приводит к изменению положения мостика (его «сгибанию»), в результате нити актина перемещаются на 1 мм к середине саркомера. Затем происходит отсоединение мостика от актина. Ритмические прикрепления и отсоединения головок миозина тянут актиновую нить к середине саркомера.
При отсутствии повторного возбуждения ионы кальция закачиваются кальциевым насосом из межфибриллярного пространства в систему саркоплазматического ретикулума. Это приводит к снижению концентрации ионов кальция и отсоединению его от тропонина. Вследствие чего тропомиозин возвращается на прежнее место и снова блокирует активные центры актина. Затем происходит фосфорилирование миозина за счет АТФ, что также способствует временному разобщению нитей. Расслабление мышцы после ее сокращения происходит пассивно – актиновые и миозиновые нити легко скользят в обратном направлении под влиянием сил упругости мышечных волокон, а также сокращения мышц-антагонистов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ЛЕКЦИЯ № 4. Физиология мышц
ЛЕКЦИЯ № 4. Физиология мышц 1. Физические и физиологические свойства скелетных, сердечной и гладких мышц По морфологическим признакам выделяют три группы мышц:1) поперечно-полосатые мышцы (скелетные мышцы);2) гладкие мышцы;3) сердечную мышцу (или миокард).Функции
4. Физиология лейкоцитов
4. Физиология лейкоцитов Лейкоциты – ядросодержащие клетки крови, размеры которых от 4 до 20 мкм. Продолжительность их жизни сильно варьируется и составляет от 4–5 до 20 дней для гранулоцитов и до 100 дней для лимфоцитов. Количество лейкоцитов в норме у мужчин и женщин
5. Физиология тромбоцитов
5. Физиология тромбоцитов Тромбоциты – безъядерные клетки крови, диаметром 1,5–3,5 мкм. Они имеют уплощенную форму, и их количество у мужчин и женщин одинаково и составляет 180–320 ? 109/л. Эти клетки образуются в красном костном мозге путем отшнуровывания от
5. Физиология фибринолиза
5. Физиология фибринолиза Система фибринолиза – ферментативная система, расщепляющая нити фибрина, которые образовались в процессе свертывания крови, на растворимые комплексы. Система фибринолиза полностью противоположна системе свертывания крови. Фибринолиз
7. Физиология всасывания
7. Физиология всасывания Всасывание – процесс переноса питательных веществ из полости желудочно-кишечного тракта во внутреннюю среду организма – кровь и лимфу. Всасывание происходит на протяжении всего желудочно-кишечного тракта, но его интенсивность неодинакова и
1. СТРОЕНИЕ И КЛАССИФИКАЦИЯ МЫШЦ. РАБОТА МЫШЦ
1. СТРОЕНИЕ И КЛАССИФИКАЦИЯ МЫШЦ. РАБОТА МЫШЦ Мышца (musculus) состоит из пучков поперечно-полосатых мышечных волокон, покрытых эндомизием (endomysium), представленным соединительнотканной оболочкой. Пучки, в свою очередь, разграничены перимизием (perimysium).Эпимизий (epimysium) покрывает
Физиология сна
Физиология сна Сон – физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим его миром. Сон является жизненно необходимым для высших животных и человека. Длительное время считали, что сон представляет собой отдых,
Упражнение 62 (для растяжения широчайших мышц спины и межреберных мышц в верхне- и среднегрудном отделах позвоночника)
Упражнение 62 (для растяжения широчайших мышц спины и межреберных мышц в верхне- и среднегрудном отделах позвоночника) Выполнять в положении сидя на стуле или стоя. Одна рука выпрямлена, поднята на уровень плеч и прижата к передней поверхности туловища, другая рука
Физиология ожирения
Физиология ожирения В животном мире нет таких форм ожирения, которые могли бы вызвать ограничения в способности передвигаться, охотиться, летать, прыгать, лазать по деревьям. Животные, у которых большие отложения жира являются видовыми признаками (киты, моржи, тюлени,
Физиология сна
Физиология сна Согласно определению специалистов, сон — это естественное физиологическое состояние человека, характеризующееся цикличностью, периодичностью, относительным уменьшением уровня физической и психической активности, отсутствием сознания и снижением
ФИЗИОЛОГИЯ ДЫХАНИЯ
ФИЗИОЛОГИЯ ДЫХАНИЯ Уважаемый читатель, давайте сделаем очень лаконичный экскурс в физиологию дыхания, чтобы уже затем показать значимость дыхательных тренингов для коррекции ВСД.В норме артериальная кровь содержит 95–98 % оксигемоглобина (HbO2) — соединения гемоглобина
Физиология дыхания
Физиология дыхания Дыхание – основа существования любого живого организма. При дыхании тело поглощает кислород из окружающей среды и выводит из организма углекислый газ – «отработанное топливо» биохимических реакций. Кислород мы расходуем практически для всех
УПРАЖНЕНИЯ ДЛЯ РАЗВИТИЯ ГРУДНЫХ МЫШЦ И МЫШЦ СПИНЫ
УПРАЖНЕНИЯ ДЛЯ РАЗВИТИЯ ГРУДНЫХ МЫШЦ И МЫШЦ СПИНЫ Развивать все мышцы тела надо пропорционально, гармонично. Чрезмерное развитие грудных мышц может привести к сутулости, поэтому упражнения для грудных мышц надо обязательно сочетать с упражнениями для мышц спины. Для
УПРАЖНЕНИЯ ДЛЯ РАЗВИТИЯ ДВУГЛАВЫХ МЫШЦ ПЛЕЧА УПРАЖНЕНИЯ ДЛЯ РАЗВИТИЯ МЫШЦ РУК И ПЛЕЧЕВОГО ПОЯСА
УПРАЖНЕНИЯ ДЛЯ РАЗВИТИЯ ДВУГЛАВЫХ МЫШЦ ПЛЕЧА УПРАЖНЕНИЯ ДЛЯ РАЗВИТИЯ МЫШЦ РУК И ПЛЕЧЕВОГО ПОЯСА Первое время часть упражнений лучше включать в утреннюю зарядку, а затем уже можно отводить для всего комплекса целиком специальное время. Каждое упражнение повторяйте 15—20
8.2. Физиология
8.2. Физиология Новости эти я знал с детства: одна страна угрожает другой, кто-то кого-то предал, экономика переживает упадок, Израиль и Палестина за протёкшие пятьдесят лет так и не пришли к соглашению, ещё один взрыв, ещё один ураган оставил тысячи людей без крова. Паоло