Глава X
Глава X
БИОЛОГИЧЕСКИЕ ПОТРЕБНОСТИ И ВИСЦЕРАЛЬНЫЕ ОЩУЩЕНИЯ
ГОЛОД И ЖАЖДА
Наиболее простой способ описания участия мозга в формировании простейших субъективных переживаний и ощущений, возможно, состоит в том, чтобы проследить, как были собраны экспериментальные данные, относящиеся к определенным ощущениям, например таким, как голод и жажда.
Несколько лет назад, когда думали, что физиологической основой таких ощущений являются висцеральные структуры, расположенные за пределами центральной нервной системы, изучалась гипотеза, согласно которой ощущение голода возникает при сокращениях желудка. Сокращения желудка наблюдались непосредственно при вскрытии брюшной полости у животных или ва время операций у человека. Сокращения желудка измерялись также косвенным путем: человек заглатывал баллон с выкаченным воздухом, который наполнялся им, когда попадал в желудок. Баллон соединялся с помощью трубки с измерительным прибором. Таким путем были установлены корреляции между сокращениями желудка и сообщениями о субъективных ощущениях голода.
Подобным образом были получены данные о том, что сухость во рту и горле коррелирует с ощущением жажды. Эти наблюдения широко известны.
Но как возникают сокращения желудка и сухость во рту? Предполагалось, что они возникают вследствие изменения состава циркулирующей крови. В экспериментах по изучению голода кровь, взятая от голодной собаки, вводилась в вену сытой собаки, у которой измерялись сокращения желудка. Эти инъекции вызывали ожидаемые сокращения, тогда как инъекция крови, взятой от сытой собаки, таких сокращений не вызывали (Luckhardt and Carlson, 1915). Повышение или понижение концентрации крови посредством внутривенной инъекции гипотонического или гипертонического водного раствора ослабляет или увеличивает ощущение жажды и устраняет или увеличивав!! сухость во рту.
Но на что действует химический или осмотический состав крови? Ответ на этот вопрос потребовал много лет исследований. Как уже отмечалось, сначала было установлено, что повреждение определенных участков ствола мозга животного приводит к появлению большого количества сахара в моче. Повреждение соседних структур ведет к ожирению. Являются ли эти центры именно тем местом, в котором голод вызывает химические изменения? Вызывают ли изменения количества сахара, циркулирующего в крови, сокращения желудка и чувство голода? Чтобы ответить на эти вопросы, необходимо было провести более точные исследования.
Исследователи соединяли молекулу сахара с другими молекулами, которые легче обнаружить при их появлении в мозгу. Было найдено вещество – золотой сероглюконат, явившееся эффективным индикатором тех областей мозга, которые избирательно поглощают молекулы глюкозы. И оказалось, что эти области в значительной степени связаны с теми, которые при их поражении вызывают ненормальное выделение сахара и расстройства питания (Mayer, 1963).
Как и в любой программе экспериментов, занимавшей научный мир в течение целых десятилетий, обнаруживались расхождения и противоречия в результатах, и многие из появившихся таким образом парадоксов, хотя и не все, можно было разрешить, лишь путем усложнения общей картины. Так, образования, ответственные за сахарный обмен, возможно, играют важную роль и в регуляции процессов питания, то есть в обмене жиров и белков, но как именно это происходит, еще предстоит изучить.
Открытие локализации рецепторных аппаратов жажды было весьма драматичным. В верхние отделы ствола мозга козы была вставлена маленькая канюля и введено небольшое количество концентрированного солевого раствора (Andersson, 1958). Коза немедленно выпила несколько литров воды. Анатомические исследования показали, что область ствола мозга, куда вводился раствор, имела огромное число сосудов и обладала особой чувствительностью к концентрации электролитов в крови.
В случае жажды большое значение в регуляции водного баланса имеют отношения между стволом мозга и гипофизом; в таких особых взаимосвязях с гипофизом находятся также многие другие ядра мозга и фактически некоторые из этих связей приводятся в действие, по-видимому, секрецией клеток мозговых ядер, поступающей в эту железу через сеть вен, связывающих гипоталамус с гипофизом (см. рис. Х-1). Кроме того, наиболее вероятно, что клетки мозга, чувствительные к этим химическим агентам и участвующие в регуляции голода и жажды, скорее распределены по системе мозгового ствола, чем сконцентрированы в одном «центре», хотя можно выделить некоторые важнейшие части данной системы.

Механизм, посредством которого рецепторные клетки осуществляют свое регулирующее действие, оказывается значительно более интересным, чем это предполагалось вначале. Как уже отмечалось, разрушение вентромедиальной области гипоталамуса ведет к ожирению. Крысы с таким поражением мозга непрерывно едят. Кажется, что, начав есть, они не в состоянии остановиться. (Они также много пьют, но это может быть связано с потребностью сделать пищу более жидкой, чтобы облегчить пищеварение.)
Таким образом, разрушенные области ответственны не столько за голод, сколько за насыщение организма. И действительно, когда с помощью глубинных электродов была осуществлена регистрация электрической активности этого центра «насыщения», было обнаружено, что его клетки активны при большой разнице в количестве сахара, циркулирующего в артериях и венах и питающего мозг; эти клетки не активны, когда это различие в концентрации сахара невелико (см. рис. Х-4, Х-5).
МЕХАНИЗМЫ ЗАПУСКА П ТОРМОЖЕНИЯ ЭЛЕМЕНТАРНЫХ ПОТРЕБНОСТЕЙ
Однако какими бы определенными ни были эти результаты, они оставляют нерешенным первоначальный вопрос – вопрос о том, как возникает голод. Эта проблема оставалась нерешенной до тех пор, пока не были проведены некоторые эксперименты. О;;;1 внесли известную ясность в этот вопрос.
Я занимался изучением функций не ствола мозга, а височной доли полушарий и обнаружил, что расстройства в акте еды (рис. Х-2), возникающие при удалении значительной части этой доли, могут быть вызваны также удалением небольшой части медиальной поверхности ее полюса – амигдалы, структуры лим-бической системы, относимой обычно к одному из базальных ганглиев (рис. Х-3). Мои опыты были проведены на обезьянах, пищевое поведение которых особенно трудно для изучения. Обезьяны сохраняют пищу в защечных мешках; они очищают и растирают предложенную им пищу, пробуют и выбрасывают ее, когда насытятся. И все же предварительные количественные результаты показали, что удаление амигдалы удваивает количество поглощенной пищи.

Рис Х-2 Влияние двустороннего удаления амигдалы на поглощение пищи обезьяной, содержащейся на свободной диете Стрелки обозначают операцию резекцию первой, а затем второй амигдалы (Pribram and Bagshow, 1953)

Рис Х-3 Реконструкция системы связей между лимбическими (амигдала и гиппокамп) и стволовыми (гипоталамус) структурами мозга (Krieg, 1966).
Мои коллеги по отделению физиологии в Йельском университете, которые научились вызывать ожирение у крыс, разрушая гипоталамический механизм «насыщения», и особенно Дж. Бробек заинтересовались этой проблемой и вместе с Б. Анандом, ординатором из Индии, начали над нею работать. Мы спланировали свои действия: я должен был производить хирургические повреждения, тогда как упомянутые сотрудники вводили в амигдалу зонд для разрушения ее электрическим током Мы создавали такие условия, которые, как мы полагали, должны были дать ответ на вопрос, связано ли расстройство питания с изменением вкусовых ощущений (последнее измерялось путем относительного поглощения горького и сладкого растворов и смеси из опилок и колесной мази и жирного пюре, по виду и консистенции похожего на эту смесь, но вполне съедобного для животных) или с некоторым нарушением обмена веществ (у животных регулярно измерялась температура и вес, а также двигательная активность животного, помещенного в специальное колесо).
Нашим руководителем, благословившим эти исследования, должен был стать Р. Берне. Все наши тщательные и кропотливые измерения оказались напрасными: результаты опытов были драматичны и видны невооруженным глазом Прежде всего половина крыс погибла не от операции, а до нее, потому что они пили очень много горького раствора и ели так много тавота и опилок, что серьезно заболели, получив хинное отравление. Дикие крысы, как известно, различают, что им полезно (и те из крыс, которые оставались в живых, также различали это, потому что они никогда не прикасались снова к вредным веществам), но специально выведенные в лаборатории белые крысы не обладали такой способностью И хотя я действительно обнаружил, что после удаления амигдалы крысы снова начинают пить горький раствор и поглощать опилки с тавотом, у меня не было ощущения, что эксперимент завершен А поскольку этот опыт также не дал ответа на вопросы, относящиеся к изучаемой проблеме, мы никогда не сообщали о его результатах.
С моими коллегами, Анандом и Бробеком, случилось даже худшее или, возможно, лучшее в зависимости от того, как на это посмотреть Почти все их крысы погибали после стереотаксической операции – и снова не от самого хирургического вмешательства, а от того, что животные прекращали одновременно есть и пить. Эти результаты были противоположны тем, которые я получил на обезьянах, хотя временный послеоперационный отказ от пищи и питья часто имел место и у них. На этой стадии почти год спустя после начала экспериментов мы, в общем, не могли считать полученные результаты удовлетворительными. Поэтому Ананд (который вскоре должен был вернуться в Индию) взял на себя тяжелый труд провести анатомический анализ разрушений мозга, которые произвели он и Бробек.
И вот новый сюрприз. Стереотаксический прибор произвел разрушение совсем не в том месте, куда он был направлен, а именно он разрушил не амигдалу, а образование, расположенное гораздо ближе к средней линии мозга – между амигдалой и гипоталамусом (рис. Х-4).

Рис. Х-4. Схема среза, проходящего через гипоталамус крысы, дающая представление о локализации в латеральном гипоталамусе «центра голода». Черным показаны области разрушения (Anand and Brobeck, 1951).
Таким образом, в латеральном гипоталамусе был открыт механизм голода, или центр Ананда – Бробека (Anand and Brobeck, 1955). Ананд, на протяжении ряда лет проводивший исследование этого вопроса, показал, что механизмы, побуждающие к еде, и механизмы насыщения находятся в реципрокных отношениях: когда центр насыщения при регистрации с помощью микроэлектродов обнаруживает активность, область, побуждающая к еде, тормозится. Когда же механизм насыщения находится в состояни покоя (как в случае низкого уровня сахара в крови), центр голода растормаживается и становится активным (Anand, 1963; рис. Х-5).
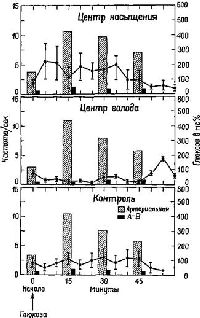
Рис. Х-5. График отдельно показывает среднюю частоту, нейронных разрядов, записанных от центра насыщения к центра голода, а также от контрольных областей. На рисунке представлена корреляция нейронных разрядов с уровнем глюкозы в артериальной крови и количеством усвоенной глюкозы (различие между артериальной и венозной концентрацией, А-В). По-видимому, изменение частоты нейронных разрядов от центра насыщения и центра голода лучше коррелирует с количеством усвоенной глюкозы, чем с ее абсолютным уровнем в артериальной; крови (см. Anand, 1963).
Оказалось, что центр голода и жажды в латеральном гипоталамусе состоит не из массы клеток, рецепторов, чувствительных к физико-химическому раздражению. Скорее, он представляет собой место пересечения путей от различных частей мозга, связывающих периферические и центральные станции, имеющие отношение к началу и прекращению еды и питья. Детали организации этой системы еще пока не установлены. Как уже указывалось, многое известно о взаимоотношении между ощущением голода и жажды и такими процессами, как сокращение и наполнение желудка, сухость во рту и т. д., которые составляют периферическую часть системы. Значительно меньше известно о центральных механизмах этой системы. Но начало уже положено (Brobeck, 1963).
НЕКОТОРЫЕ ФАКТЫ
Амигдала, с изучения которой начались наши эксперименты, является одной из структур, откуда идут волокна в латеральную гипоталамическую область пересечения нервных путей. Ее функции в регуляции питания и питья продолжают тщательно изучаться. Хотя инъекции малейших доз некоторых химических агентов (оказывающих адренэргическое или холинэргическое действие) в гипоталамический центр насыщения могут вызвать или остановить еду и питье (в зависимости от характера химического агента), такие же инъекции в амигдалу никакого эффекта не вызывают. Но если еда или питье уже начались, введение тех же самых химических агентов может изменить количество съеденной пищи или выпитой воды (Grossman, 1966; рис. Х-6Б). Были установлены ясные количественные соотношения между дозой вводимого вещества и количеством поглощенной воды (Russel et al., 1968; рис. Х-6А), а кривая, характеризующая эти соотношения, напоминает многие кривые порогов, установленные в психофизических экспериментах, проведенных со зрительной и слуховой модальностями. Таким образом, психологические процессы экстероцептивных и висцеральных ощущений имеют много сходного.

Рис. Х-6. А – отношения между изменением потребления воды и дозой карбахола (холин-эргетика), вводимого в амигдалу при трех уровнях водной депривации. Чтобы избежать отрицательных логарифмов, шкала по абсциссе представлена в молях ХЮ~" (RusselJ et ai., 1968);

Б – влияние адренэргической и холинэргической стимуляции гипоталамуса у сытых крыс на скорость нажатий на педаль для получения пищевого и водного подкрепления по программе с варьирующим около 30 сек интервалом. Контрольные уровни определялись в течение одночасового периода перед каждой стимуляцией (см, Grossman1962.)
Это совпадение, вероятно, не случайно. Однажды мне довелось обследовать некоторых больных с двусторонним удалением медиальной части височного полюса, включая амигдалу. Эти больные, так же как их двойники среди обезьян, ели значительно больше, чем нормальные люди, и их вес превышал нормальный на сто фунтов. Наконец, я спросил одного такого больного, как же он себя чувствует, постоянно ощущая голод. Но к моему большому удивлению, ожидаемого ответа не последовало. Одна больная, которая после операции прибавила за год в весе более чем сто фунтов ( 45 кг ), была обследована во время завтрака. Была ли она голодна? Она ответила: «Нет». Хотела ли бы юна съесть кусок мяса, сочный бифштекс? Она ответила: «Нет». А хотела ли бы она шоколадную конфету? Она хмыкнула, но, когда конфету ей не дали, больше к этому предмету не возвращалась. Несколько минут спустя, когда обследование закончилось и открылись двери общей комнаты, она увидела других больных, завтракавших за длинным столом. Больная бросилась к столу, растолкала других больных и начала набивать едой рот обеими руками. Ее немедленно позвали в кабинет и повторили вопросы, относящиеся к еде. Были получены те же самые отрицательные ответы даже после многозначительного противопоставления этих ответов ее недавнему поведению за столом. Повреждение мозга каким-то образом ослабило у больной ощущение голода и насыщения, и это ослабление сопровождалось чрезмерной едой.
Пока мы плохо понимаем, как происходит такое нарушение. Тем не менее этот пример ясно показывает, как нелепо предполагать, что между определенным типом поведения и инстроспек-тивным отчетом о нем существует прямое соответствие. Должны ли мы сказать, что больная ощущала голод, потому что она жадно ела, несмотря на ее отрицание наличия голода? Или мы должны рассматривать ее утверждения как истинные и искать объяснение ее неумеренной еды в чем-то другом? Этот парадокс может быть разрешен, если, как и в предыдущих главах, посвященных экстероцептивным ощущениям, мы будем считать, что поведение складывается из нескольких процессов, одним из которых является ощущение от своего состояния, описываемое в отчете испытуемого.
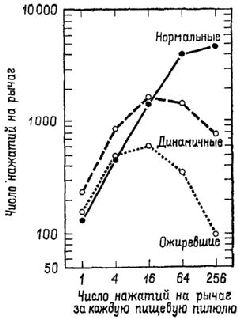
Рис. Х-7. Среднее число нажатий на рычаг (за каждые 12 чалов) у нормальных, ожиревших (поздняя стадия гиперфагии) и динамичных (до ожирения, ранняя стадия гиперфагии) животных в зависимости от числа нажатий, необходимых для получения пищевого шарика (Teitelbaum, 1957).
Аналогичный парадокс возникал перед исследователями функций гипоталамуса. Выше уже отмечалось, что, когда разрушения производились в области вентромедиального ядра гипоталамуса, оперированные крысы ели значительно больше контрольных и толстели. Но это еще не все. Хотя крысы с таким повреждением гипоталамуса ели больше, когда еда была вполне доступной, они в меньшей степени активно искали пищу, когда появлялись какие-либо препятствия (Miller, Bailey and Stevenson, 1950; см. также рис. Х-7). Было также обнаружено, что, чем вкуснее еда, тем больше ее съедают оперированные животные (Teitelbaum, 1955). Это наводит на мысль, что у оперированных животных больше не «влечение» к еде, а они более «разборчивы», нежели контрольные крысы. Результаты экспериментов, проведенных Кресном (1962) и Гроссманом (1966), усугубили парадокс: электрическое раздражение вентромедиального ядра вызывает прекращение поглощения пищи и воды у голодных крыс, а химическое раздражение холинэргического механизма вызывает топанье ногами (у песчанок, личное сообщение Глик-мана) и агрессивность, если животное рассердить (King and Hoe-bel, 1968). Гроссман коротко обобщил эти результаты в следующем утверждении: медиальный гипоталамус управляет изменениями висцеральных ощущений, а не потребностей. Но здесь мы снова сталкиваемся с нашей прежней дилеммой. Если медиальный гипоталамический механизм не связан с мотивацией, как же в таком случае возникает потребность в еде, питье и т. д.? Ответ на этот вопрос дают эксперименты. Вентромедиальная и латеральная гипоталамические области составляют две части одной" системы: латеральная часть является механизмом, «запускающим» питание (удаление ее ведет у крыс к голоданию), а медиальная часть является тормозящим механизмом (рис. Х-8). Парадокс разрешается, если предположить, что процессы, которые выключают организм из активного состояния, также генерируют соответствующие ощущения. Таким образом, становится яснее важнейшее различие между мотивацией и эмоцией: термин «мотивация» можно относить только к потребностям, «запускающим» процессам (тем, которые объединяются в латеральной гипоталамической области), а термин «эмоция» – к действию аффективных, «останавливающих» или насыщающих процессов, восстанавливающих равновесие.

Рис. Х-8. Механизмы «остановки» и «запуска» (Stop and Go) в терминах схемы Т-О -Т -Е.
Многие из этих положений развиваются в следующей главе. Здесь же отметим только, что объяснения – те объяснения, с помощью которых ученые могут осмыслить полученные данные, – сформулированы в терминах изменения висцеральных ощущений, потребностей (мотивов) и аффектов (эмоций), получаемых в результате разрушения мозга. Выводы об этих изменениях в ощущениях мы делаем на основе наблюдаемых нейропси-хологических данных, но при объяснении экспериментальных результатов следует учитывать не только поведенческие, но и многие другие (например, нейрологические) факты.
РЕЗЮМЕ
Эксперименты, проведенные для уточнения функций рецеп-торных ядер мозга, привели к целому ряду загадочных парадоксов, которые не могли быть разрешены до тех пор, пока описание результатов осуществлялось на субъективном языке ощущений. Для каждой из рецепторных функций были установлены мотива-ционный, «запускающий», и эмоциональный, «останавливающий», механизмы. «Запускающие» процессы выражаются через элементарные потребности, а процессы «остановки» – через аффекты.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 1
Глава 1 24. Сунь Бинь – знаменитый военачальник периода Борющихся государств, уроженец Государства Ци. Потомок Сунь-цзы. В молодости вместе с Пан Цзюанем (?-341 г. до н. э.) изучал военное дело. Позже Вэйский Сюань-ван назначил Пан Цзюаня главнокомандующим государства Вэй.
Глава 2
Глава 2 29. Бянь Цюэ – врач государства Цинь эпохи Весен и Осеней, уроженец области Бохай. В молодом возрасте работал на должности смотрителя постоялого двора. Однажды на постоялый двор заехал знаменитый врач Чжан Санцзюнь, Бянь Цюэ отнесся к нему с большим почтением и на
Глава 3
Глава 3 31 . «Книга перемен», известная также как «И цзин» – наиболее авторитетное произведение китайской канонической и философской литературы, оказавшее фундаментальное воздействие на всю культуру традиционного Китая, стран Дальнего Востока и Юго-Восточной Азии.32.
Глава 19
Глава 19 64. Цит. по: У-цзин. Семь военных канонов Древнего Китая, – СПб.: Евразия, 1998, стр. 155.65. Сунь Икуй – врач династии Мин, уроженец уезда Сюйнин (ныне уезд Сюйнин в провинции Аньхуй). Точные годы жизни и смерти неизвестны, но есть свидетельства, что его деятельность
Глава 4 Чуа
Глава 4 Чуа Моя фамилия - Чуа (Кай - на мандаринском), и я люблю её. Мои близкие - выходцы из южной китайской провинции Фуцзянь, известной своими филологами и учёными. Один из моих предков со стороны отца, всесторонне образованный Чуа By Ненг, был астрономом императора Шэнь
Глава 11
Глава 11 Если рождение – начало естественной жизни человека, то смерть – естественный итог ее завершения. Петровича давно интересовала тема смерти. Ее неизбежность, таинственность, загадочность и некий суеверный страх. Как-то выбрав время, он решил расспросить об этом
Глава 12
Глава 12 Человек не должен заниматься своим здоровьем, это должно быть его естественным состоянием, как результат естественной жизни. – Послушай, Гея, а как у вас дела обстоят с лечением? – спросил Петрович.– Болезней как таковых у нас нет. Чаще всего у нас бывают ушибы,
Глава 2
Глава 2 1. Weinberg, Robert A., 1996. How Cancer Arises. Scientific American, Special Issue, What you need to know about Cancer, September, 275, 3, 62–70.2. Pollack, M.N. and others, 1998. IGF-1 Risk Factor for Prostate Cancer. Science, 279, 563–566.Pollack, M.N., Huynh, H.T., and Lefebvre, S.P., 1992. Tamoxifen reduces serum insulin-like growth factor 1 (IGF-1). Br Cancer Res Treat, 22, 91-100.Pollack, M.N., Polychronakos, C., Yousefi, S., and Richard, M., 1988. Characterisation of
Глава 3
Глава 3 1. Trichopoulos, Dimitros, Li, Frederick, V. and Hunter, David J., 1996. What Causes Cancer? Scientific American, Special Issue, What you need to know about Cancer. September 275, 3, 80–85.2. Boots the Chemist, 1998. Breast Awareness – an essential guide, Be enlightened, not frightened. Published in association with the charity Breast Cancer Care.3. Trichopoulos, Dimitrios, Li, Frederick, P. and Hunter, David J., 1996. What Causes Cancer Scientific American, Special Issue,
Глава 4
Глава 4 1. Сох, Peter and Brusseau, Peggy, 1995. LifePoints For Kids. Bloomsbury.2. Statistical Abstracts of the US, 1994 edition.3. Willett, WC. and others, 1996. Strategies for Minimizing Cancer Risk. Scientific American, Special Issue, What you need to know about Cancer, 275:3, pp, 88–95.4. Akre, James, ed., 1989. Infant feeding: the physiological basis. WHO Bulletin, OMS Supplement, Vol. 67, and Lawrence, Ruth A., 1994. Breastfeeding: A guide for the medical profession.
Глава 6
Глава 6 1. Gerhard Schrauzer, Ph.D., of the University of California cited in Better Nutrition, Nov, 1989, v 51, no. 11, p. 14 (2).2. Redhook., April, 1989, v 172, n6, p. 96 (5).3. Spallholz, J.E., Stewart, J.R., 1989. Advances in the role of minerals in immunobiology. Center for Food and Nutrition, Texas Tech University, Lubbock 79409. Biol Trace Elem Res Mar, 19 (3) pp. 129–151.4. Archives of Environmental Health, September/October 1976.5. Knekt, Paul, and others, 1988. Selenium
Глава 7
Глава 7 1. Campbell, T.C. and Junshi, C., 1994. Diet and chronic degenerative disease perspectives from China. Am. J Clin. Nutr, 59, Suppl., 11 535-11 615.2. Packham, D.E., 1999. Impacts of Commercialisation and Privatisation on Capabilities for Scientific Advice, Oracles or Scapegoats? IPMS Conference notes. October.3. Packham, D.E., 1999. Impacts of Commercialisation and Privatisation on Capabilities for Scientific Advice, Oracles or Scapegoats? IPMS Conference notes,
Глава 22
Глава 22 Дженн и Билли познакомились летом 2002 года, когда Билли, окончив первый курс Университета штата Виргиния, вернулся домой работать на спасательной станции в городе Виргиния-Бич. Однажды утром он прибыл на свой пост и обнаружил, что поговорка «Дураку — счастье»
Глава 23
Глава 23 Мы прибыли в Крил, когда стемнело. Автобусик, дребезжа, дотащился до остановки, взвизгнув напоследок тормозами, словно издал вздох облегчения. За окном я разглядел призрачную старую соломенную шляпу Кабальо. Подпрыгивая, она плыла в темноте в нашу сторону.Мне