Мембранные органеллы. Транспорт через мембраны
Мембранные органеллы. Транспорт через мембраны
Для клеток человека характерно наличие огромного количества внутриклеточных мембран, образующих несколько компартментов (от англ. compartment – «отделение, купе»), отличающихся друг от друга строением и функцией: цитозоль, ядро, эндоплазматический ретикулум, комплекс Гольджи, митохондрии, лизосомы, пероксисомы. Благодаря наличию этих элементов в клетке одновременно протекает большое количество различных биохимических реакций.
Все мембранные органеллы построены из элементарных мембран, принцип строения которых аналогичен описанному выше строению плазмолеммы. Поглощение клетками макромолекул и частиц происходит путем эндоцитоза (от греч. endon – «внутри», kytos – «клетка»), выделение – путем экзоцитоза (от греч. ехо – «вне», kytos – «клетка»).
Одной из важнейших функций плазмолеммы является транспорт. Напомним, что обращенные друг к другу гидрофобные «хвосты» липидов препятствуют проникновению полярных водорастворимых молекул. Различают два вида транспорта: пассивный и активный. Первый не требует затрат энергии, второй энергозависимый. Как правило, внутренняя (цитоплазматическая) поверхность мембраны несет отрицательный заряд, что облегчает проникновение в клетку положительно заряженных ионов. Вода поступает в клетку путем осмоса (от греч. osmos – «толчок, давление»), который представляет собой медленное проникновение воды через полупроницаемую мембрану, разделяющую два раствора различной концентрации. В результате концентрация этих двух растворов выравнивается.
Диффузия (от лат. diffusion – «распространение, растекание») – это переход ионов или молекул, вызванный их броуновским движением через мембраны из зоны, где эти вещества находятся в более высокой концентрации, в зону с более низкой концентрацией до тех пор, пока концентрации по обе стороны мембраны выравниваются. Специфические транспортные белки, встроенные в мембрану, переносят через нее небольшие полярные молекулы, причем каждый белок осуществляет транспорт одного класса молекул или только одного соединения. Некоторые трансмембранные белки образуют каналы. Активный транспорт осуществляют белки-переносчики, при этом расходуется энергия, получаемая вследствие гидролиза АТР (аденозинтрифосфатной кислоты) или протонного потенциала. Активный транспорт происходит против градиента концентрации. Для осуществления биохимических реакций необходимо поступление веществ в клетку путем эндоцитоза и выведение продуктов обмена экзоцитоза.
Эндоцитоз. Существует несколько способов эндоцитоза. Поступление жидких коллоидных частиц называется пиноцитозом, а крупных твердых частиц – фагоцитозом. Для того чтобы внешние молекулы поступили в клетку, они должны быть сначала связаны рецепторами гликокаликса. Цитолемма начинает впячиваться, затем ее края сближаются и смыкаются, отщепляя пузырек, несущий в себе захваченные молекулы. Образуется эндреома, которая погружается в цитоплазму и встречается с лизосомами. Их мембраны сливаются. В возникшей вторичной лизосоме вещества, поступившие в клетку, подвергаются расщеплению.
Экзоцитоз обеспечивает выведение крупномолекулярных соединений. Сначала они сегрегируются в комплексе Гольджи в виде транспортных пузырьков и направляются к клеточной поверхности. Мембрана пузырька встраивается в цитолемму, и содержимое пузырька оказывается за пределами клетки.
Известны две разновидности эндоцитоза: фагоцитоз – поглощение частиц (от греч. phagos – «пожирающий» и kytos – «клетка») и пиноцитоз – поглощение растворенных веществ (от греч. Pino – «пью»). Фагоцитированная частичка, заключенная в мембрану, называется фагосомой. В процессе эндо– и экзоцитоза транспортируемые вещества заключены в мембранные пузырьки.
Эндоплазматическая сеть, или эндоплазматический ретикулум (ЭР), представляет собой единую непрерывную полость, ограниченную мембраной, образующей множество инвагинаций и складок (см. рис. 1). Поэтому на электронограммах эндоплазматическая сеть выглядит в виде множества трубочек, плоских или округлых цистерн, мембранных пузырьков. Различают два типа ЭР: гранулярный и агранулярный. Обращенная к цитозолю сторона первого покрыта рибосомами, второго лишена их. Функция гранулярного ЭР: синтез белков рибосомами и транспорт белков, гладкого синтез и обмен углеводов и липидов (стероидных гормонов, гликогена, холестерола) и обезвреживание (гепатоциты), синтез хлоридов, из которых в желудке образуется соляная кислота. Будучи депо ионов кальция, гладкий ЭР участвует в мышечном сокращении; отграничивает будущие тромбоциты в мегакариоцитах. Одной из важнейших функций ЭР является синтез мембранных белков и липидов для всех клеточных органелл.
Комплекс, или аппарат Гольджи (КГ), представляет собой совокупность цистерн, пузырьков, пластинок, трубочек, мешочков, ограниченных мембраной, в которых накапливаются и упаковываются синтезированные продукты (см. рис. 1). Эти продукты с помощью элементов комплекса выводятся из клетки, кроме того, в них происходит синтез полисахаридов, образование белково-углеводных комплексов и модификация переносимых молекул. В световом микроскопе КГ выглядит в виде сеточки или системы канальцев и вакуолей. КГ имеется во всех клетках человека, кроме эритроцитов и роговых чешуек эпидермиса. В большинстве клеток КГ расположен вокруг или вблизи ядра. В КГ выявляются три мембранных элемента: уплощенные мешочки (цистерны), пузырьки и вакуоли. КГ – трехмерная структура чашеобразной формы, состоящая из нескольких (от одной до нескольких сот) диктиосом (от греч. dyktion – «сеть»). Каждая диктиосома содержит 4–8 (в среднем 6) лежащих параллельно уплощенных цистерн, пронизанных порами с расширенными концами, от которых отщепляются вакуоли, содержащие синтезированные вещества. Цистерны ассоциированы со множеством мембранных пузырьков, а также с более крупными секреторными гранулами. Элементы комплекса Гольджи связаны между собой каналами.
Мембраны комплекса Гольджи образуются и поддерживаются гранулярным эндоплазматическим ретикулумом, в котором синтезируются мембранные компоненты. Они переносятся транспортными пузырьками, отпочковывающимися от ЭР, и сливаются с КГ, от которого постоянно отпочковываются секреторные пузырьки, а мембраны цистерн постоянно обновляются. Они поставляют гликокаликс и синтезированные вещества к плазмолемме, таким образом обеспечивается и обновление плазмолеммы. Одной из важнейших функций КГ является сортировка белков.
Лизосомы – мембранные органеллы, содержащие около 50 видов различных гидролитических ферментов, которые синтезируются на рибосомах гранулярного эндоплазматического ретикулума, откуда переносятся транспортными пузырьками в КГ, где они видоизменяются. От поверхности КГ отпочковываются первичные лизосомы. Все лизосомы клетки формируют единое лизосомное пространство, в котором постоянно поддерживается кислая среда рН колеблется в пределах 3,5–5,0. Мембраны лизосом устойчивы к заключенным в них ферментам и предохраняют цитоплазму от их действия.
Различают четыре функциональные формы лизосом. Первичные лизосомы, отпочковавшиеся от комплекса Гольджи, сливаясь с фагосомой, образуют вторичную лизосому (фаголизосому), в которой происходит переваривание поглощенных веществ до мономеров. Последние транспортируются через лизосомальную мембрану в цитозоль. Непереваренные вещества остаются в лизосоме, в результате чего образуется остаточное тельце. Кроме того, лизосомы переваривают поврежденные структуры собственной клетки (аутолизосома).
Пероксисомы представляют собой пузырьки диаметром от 0,2 до 0,5 мкм, окруженные мембранами, содержащие окислительные ферменты (около 40 % всех белков составляет каталаза), производящие и разрушающие перекись водорода. Они используют молекулярный кислород.
Митохондрии, являющиеся «энергетическими станциями клетки», участвуют в процессах клеточного дыхания и преобразования энергии в форму, доступную для использования клеткой. В световом микроскопе митохондрии выглядят в виде округлых, удлиненных или палочковидных структур длиной 0,3–5,0 мкм и шириной 0,2–1,0 мкм. Количество, размеры и расположение митохондрий зависят от функции клетки, ее потребности в энергии. Так, в каждой печеночной клетке их количество достигает 2500. С помощью электронной микроскопии установлено, что митохондрии являются органеллами с двойными мембранами (рис. 5). Между наружной и внутренней митохондриальными мембранами расположено межмембранное пространство. Внутренняя мембрана образует многочисленные складки, или кристы, благодаря которым внутренней мембраны резко возрастает. На внутренней поверхности крист лежит множество электронноплотных субмитохондриальных элементарных частиц (до 4000 на 1 мкм2 мембраны), имеющих форму гриба. В пространстве, ограниченном внутренней митохондриальной мембраной, находится мелкозернистый матрикс.
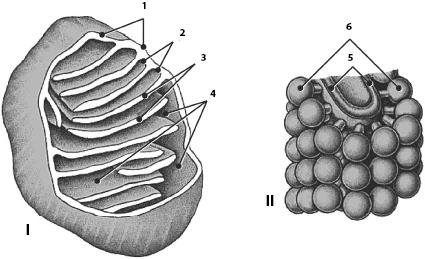
Рис. 5. Митохондрия (по Б. Албертсу и др.; по К. де Дюву, с изм.). I – общая схема строения: 1 – наружная мембрана; 2 – внутренняя мембрана; 3 – кристы; 4 – матрикс; II – схема строения кристы: 5 – складка внутренней мембраны; 6 – грибовидные тельца
Митохондрии содержат собственную ДНК, РНК и рибосомы, которые находятся в матриксе. Таким образом, митохондрии снабжены собственной генетической системой, необходимой для их самовоспроизведения и синтеза белков. Следует подчеркнуть, что митохондриальные ДНК, РНК и рибосомы отличаются от таковых собственной клетки и весьма сходны с прокариотическими.
ВНИМАНИЕ
У млекопитающих, в том числе и человека, митохондриальный геном наследуется от матери.
Митохондрии размножаются путем деления уже существующих независимо от деления других митохондрий и самой клетки.
В клетках постоянно происходит метаболизм (от греч. metabole – «перемена, превращение»), или обмен веществ, который представляет собой совокупность процессов ассимиляции (реакции биосинтеза сложных биологических молекул из более простых) и диссимиляции (реакции расщепления). В результате диссимиляции освобождается энергия, заключенная в химических связях веществ. Эта энергия используется клеткой для осуществления различной работы, в том числе и ассимиляции. Напомним, что энергия не возникает и не уничтожается, она лишь переходит из одного вида в другой, пригодный для выполнения работы. Клетка использует энергию, заключенную в химических связях аминокислот, моносахаридов и жирных кислот. Они образуются в результате пищеварения из белков, углеводов и жиров и поступают в клетку.
Рассмотрим энергетический обмен на примере расщепления глюкозы. Глюкоза транспортируется через плазматическую мембрану, и в цитоплазме происходит ее бескислородное расщепление, или гликолиз. Гликолиз – это многоступенчатый ферментативный процесс, в результате которого из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты и две молекулы АТР (с учетом двух молекул АТР, затрачиваемых для осуществления реакций). Пировиноградная кислота подвергается дальнейшему окислению (аэробному при участии кислорода) в митохондриях, в которых имеются цепи ферментов, катализирующие реакции синтеза АТР (аденозинтрифосфат). АТР является универсальным переносчиком и основным аккумулятором энергии в клетке. Энергия заключена в высокоэнергетических связях между остатками фосфорной кислоты.
При отщеплении от АТР одной фосфатной группы образуются АДР (аденозиндифосфатная кислота) и фосфат и выделяется свободная энергия, которая используется клеткой для осуществления работы. В митохондриях АДР, соединяясь с остатком фосфорной кислоты, превращается в АТР. В результате гликолиза освобождается лишь около 5 % энергии, остальная освобождается в митохондриях в процессе аэробного окисления и запасается в АТР. В расчете на одну молекулу глюкозы образуется 36 молекул АТР.
Ядро – основная клеточная структура, имеется во всех клетках человека, кроме эритроцитов и тромбоцитов. В большинстве клеток его форма шаровидная или овоидная, однако встречаются и другие формы ядра (кольцевидное, палочковидное, веретеновидное, четковидное, бобовидное, сегментированное, полиморфное и др.). Размеры ядер колеблются в широких пределах от 3 до 25 мкм. Наиболее крупное ядро имеет яйцеклетка. Большинство клеток человека одноядерные, однако имеются двухъядерные (например, некоторые нейроны, гепатоциты, кардиомиоциты), а некоторые структуры многоядерные (мышечные волокна миосимпласты).
В ядре различают следующие структуры: ядерную оболочку, хроматин, ядрышко и нуклеоплазму. Ядро окружено ядерной оболочкой, состоящей из внутренней и наружной ядерных мембран толщиной 8 нм каждая, разделенных перинуклеарным пространством (или цистерной ядерной оболочки) шириной 20–50 нм. Обе являются элементарными клеточными мембранами. К наружной, переходящей в гранулярный эндоплазматический ретикулум, прикреплены рибосомы. Перинуклеарное пространство составляет единую полость с эндоплазматическим ретикулумом (см. рис. 1).
Ядерная оболочка пронизана множеством расположенных упорядоченно ядерных пор округлой формы диаметром 50–70 нм, которые в общей сложности занимают до 25 % поверхности ядра. Через ядерные поры осуществляется избирательный транспорт крупных частиц, а также обмен веществ между ядром и цитозолем.
В живых клетках кариоплазма (нуклеоплазма) гомогенна (кроме ядрышка). После фиксации и обработки тканей для световой или электронной микроскопии в ней становятся видными два типа хроматина (от греч. chroma – «краска»); хорошо окрашивающийся гетерохроматин (неактивный) и светлый эухроматин (активный).
В делящемся ядре хроматин спирализуется, образуя хромосомы. Хроматин неделящегося ядра и хромосомы делящегося образованы дезоксирибонуклеиновой кислотой (ДНК), связанной с РНК и белками (гистонами и негистоновыми). Следует подчеркнуть химическое тождество хроматина и хромосом.
В соматических клетках имеются по две копии каждой хромосомы, их называют гомологичными. Они одинаковы по длине, форме, строению, расположению полос и несут одни и те же гены, которые локализованы одинаково. Нормальный кариотип человека (от греч. karyon – «ядро ореха», typos – «образец») включает 22 пары аутосом и одну пару половых хромосом (ХХ женщины или XV мужчины) (рис. 6).
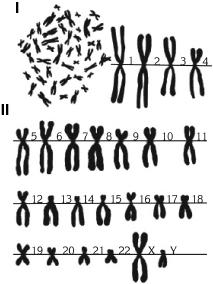
Рис. 6. Кариотип человека (здорового мужчины). I – кариотип, общий вид; II – метафазные хромосомы
Ген участок ДНК, характеризующийся определенной последовательностью нуклеотидов, ответственный за синтез специфического белка. Ген является элементарной единицей наследственности.
Ядрышко (одно или несколько) выявляется во всех неделящихся ядрах в виде плотного интенсивно окрашивающего округлого однородного базофильного тельца, величина которого пропорциональна интенсивности белкового синтеза. В ядрышке образуются рибосомы. Ядерный сок – не окрашивающаяся электронносветлая часть ядра – коллоидный раствор белков, окружающий хроматин и ядрышко.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
ТРАНСПОРТ
ТРАНСПОРТ КАКОЙ БЫВАЕТ ТРАНСПОРТ Посмотрите, самолетОтправляется в полет.Самолет летит, гудит,Смелый летчик в нем сидит.Прикусить зубами широкий кончик языка и длительно произносить звук [Л], при этом поднять руки в стороны и покачивать ими, как крыльями.Ну, а это
КАКОЙ БЫВАЕТ ТРАНСПОРТ
КАКОЙ БЫВАЕТ ТРАНСПОРТ Посмотрите, самолетОтправляется в полет.Самолет летит, гудит,Смелый летчик в нем сидит.Прикусить зубами широкий кончик языка и длительно произносить звук [Л], при этом поднять руки в стороны и покачивать ими, как крыльями.Ну, а это парашют.Вниз нас
1. Введение в курс гистологии. Органеллы клетки
1. Введение в курс гистологии. Органеллы клетки Гистология – наука о строении, развитии и жизнедеятельности тканей живых организмов. Следовательно, гистология изучает один из уровней организации живой материи – тканевый.Основным объектом гистологии в системе
4. Морфология и функции органеллы клетки (продолжение)
4. Морфология и функции органеллы клетки (продолжение) Лизосомы – наиболее мелкие органеллы цитоплазмы, представляют собой тельца, ограниченные билипидной мембраной.Функция лизосом – обеспечение внутриклеточного пищеварения, т. е. расщепление как экзогенных, так и
Газообмен и транспорт газов
Газообмен и транспорт газов Газообмен О2 и СО2 через альвеолярно-капиллярную мембрану происходит с помощью диффузии, которая осуществляется в два этапа. На первом этапе диффузионный перенос газов происходит через аэрогематический барьер, на втором – происходит
УКРЕПЛЯЕМ МЕМБРАНЫ КЛЕТОК ТРЕНИРОВКАМИ
УКРЕПЛЯЕМ МЕМБРАНЫ КЛЕТОК ТРЕНИРОВКАМИ О пользе физических упражнений сказано много. Регулярные занятия спортом улучшают работу сосудистой системы, нормализуют давление и уровень сахара, наращивают мышечную массу, сжигают жир. Именно при тренировках изменяется наш
Мембраны
Мембраны Жизнь — это вечное движение жидкостей между клетками и внутри клеток. Остановка этого движения приводит к смерти. Частичное замедление движения жидкостей в каком-то органе вызывает частичное расстройство. Общее замедление вне- и внутриклеточных жидкостей в
Транспорт
Транспорт Кровь, лимфа и спинномозговая жидкость являются транспортной системой организма. Они доставляют к клеткам необходимые питательные вещества и выводят образовавшиеся в процессе жизнедеятельности шлаки и токсины. Как только процесс выведения этих отходов по
Через сто лет я молодой 20-летний
Через сто лет я молодой 20-летний Я – Дух Божий, по святой воле Бога постоянно здоровею-крепну, постоянно здоровеют-крепнут нервы, постоянно развиваюсь, ускоряю Богом данное постоянное исцеление-омоложение, всегда становлюсь 20-летним.Я и через пять лет 20-летний. Я и через
Глава 3. Транспорт газов кровью
Глава 3. Транспорт газов кровью «Переносчиком» кислорода от легких к тканям и органам и углекислого газа от тканей и органов к легким является кровь. В свободном (растворенном) состоянии переносится настолько малое количество газов, что им можно смело пренебречь при
Вдох – через нос, выдох – через рот
Вдох – через нос, выдох – через рот Научитесь дышать брюшным дыханием так, чтобы вдыхать носом, а выдыхать ртом (рот при этом лишь чуть приоткрыт, губы сложены наподобие маленькой трубочки) – мягко, плавно, без
Глава VI ДЫХАНИЕ ЧЕРЕЗ НОЗДРИ И ДЫХАНИЕ ЧЕРЕЗ РОТ
Глава VI ДЫХАНИЕ ЧЕРЕЗ НОЗДРИ И ДЫХАНИЕ ЧЕРЕЗ РОТ Один из первых уроков науки дыхания йогов посвящается тому, чтобы научиться дышать носом и победить обычную привычку – дышать ртом.Дыхательный механизм человека позволяет ему дышать и носом и ртом, но для него дело истинно
Блок бросков через голову (броски через голову)
Блок бросков через голову (броски через голову) Бросок через голову — перебрасывание гири через голову с последующим её приёмом той же рукой с поворотом туловища на 180°. Все броски через голову выполняются с поворотом туловища на 180°, поэтому при названии броска поворот