К характеристике изменения ЭЭГ у больных, находящихся на длительном лечебном голодании В. В. АРШАВСКИЙ, Б. В. КРАЙЦЕРОВ (Москва)
К характеристике изменения ЭЭГ у больных, находящихся на длительном лечебном голодании
В. В. АРШАВСКИЙ, Б. В. КРАЙЦЕРОВ (Москва)
Из многочисленных данных литературы известно, что центральный отдел висцерального анализатора как у животных, так и у человека проецируется в передних отделах коры больших полушарий головного мозга (1, 2, 6, 11, 12, 16, 17, 19).
Было установлено, что на проведение висцеральных импульсов к коре большое влияние оказывают неспецифические структуры головного мозга — ретикулярная формация среднего мозга и моста и древние корковые образования — лимбическая кора и гиппокамп (1, 2, 4, 5, 13, 14).
Эти данные были подтверждены в ряде экспериментальных работ, в которых изучалось влияние голодания животных на биоэлектрическую активность мозговых структур. При непродолжительном 1—3-дневном голодании у животных в ЭЭГ отмечается развитие десинхронизации в передних отделах коры большого мозга. Эта десинхронизация связывается с развитием возбудительного процесса в задних ядрах гипоталамуса, а затем в ретикулярной формации среднего мозга (8, 10). В этих исследованиях показано, что пищевое возбуждение, определяющее пищевые мотивации животных в состоянии голода, охватывает целую систему функционально объединенных нервных элементов, находящихся в подкорковых и корковых структурах головного мозга. В механизме избирательной активации передних отделов коры у голодных животных решающая роль принадлежит восходящим влияниям из пищевых центров гипоталамуса. Эти влияния осуществляются через медиальные ядра таламуса. Избирательная активация передних отделов коры мозга у животных после кратковременного голодания не блокируется аминазином, подавляющим адренэргический субстрат ретикулярной формации, но снимается введением холинолитических веществ. Однако авторы отмечают, что наряду с восходящими активирующими влияниями на кору мозга, гипоталамические отделы пищевого центра у голодных животных оказывают активирующее влияние и на ретикулярную формацию среднего мозга. Это влияние усиливается при больших сроках голодания. При длительном голодании холинергические механизмы восходящих влияний на кору все в большей степени обогащаются адренэргическими механизмами восходящих активирующих влияний ретикулярной формации среднего мозга, которые могут быть заблокированы аминазином. Данные, полученные указанными авторами, подтверждают предположение П. К. Анохина (3) о специфическом характере множественных восходящих активирующих влияний подкорковых образований на кору большого мозга при реакциях различных биологических модальностей.
Подобных исследований на человеке не проводилось. Тем не менее внедряющийся в клинику соматических и психических болезней метод лечебного голодания настоятельно требует проведения подобного рода исследований. В книге Ю. С. Николаева (17) отмечается, что у людей, находящихся на длительном лечебном голодании, в ЭЭГ развивается десинхронизация во всех отведениях, сохраняющаяся длительное время.
МЕТОДИЧЕСКИЕ ЗАМЕЧАНИЯ
Нами исследовано в динамике проведения лечебного голодания 21 человек с различными диагнозами: шизофрения простая форма — 6, параноидная — 3, ипохондрическая — 5, циклотимия — депрессивная фаза — 2, атеросклероз сосудов головного мозга — начальная стадия — 1, остаточные явления органического поражения головного мозга с преимущественным вовлечением диэнцефальной области — 4 человека.
Запись ЭЭГ производилась до начала голодания, на 3—5 день голодания (в некоторых случаях эта запись производилась с применением аминазиновой пробы — аминазин вводился внутримышечно или в дозе 25—50 мг), на 10—12 день голодания, в последний день голодания (эта запись производилась с первым приемом пищи — стакан фруктового сока, последняя запись осуществлялась на 10—14 день питания больного. Все больные находились на лечебном голодании в течение от 17 до 35 дней (с последующей молочно-растительной диетой), в результате которого 12 больных дали хороший эффект, а для 9 больных лечение было безрезультатным.
РЕЗУЛЬТАТЫ НАБЛЮДЕНИЙ И ИХ ОБСУЖДЕНИЕ
Следует отметить, что изменения, обнаруженные нами в ЭЭГ больных, находящихся на длительном лечебном голодании, были в основном однотипны и проявлялись в виде десинхронизации, развивающейся в ЭЭГ в большей или меньшей степени, и появления быстрой острой активности в лобных (а иногда и в височных) отделах коры. Однако подобный эффект выявлялся в большей степени на ЭЭГ только в том случае, если фоновая кривая больного не была десинхронизирована или гиперсинхронизирована, т. е. когда фон ЭЭГ не был возбужден.
Эти изменения, возможно, связаны с развитием возбудительного процесса в ретикулярной формации ствола и задних ядер -гипоталамуса и могут обеспечиваться наличием «голодной» импульсации с интероцепторов желудка и кишечника. В эту стадию голодания малые дозы аминазина снимают я ЭЭГ десинхронизацию, развивающуюся особенно четко в передних отделах. Следует отметить, что у больных, которые хотели в это время есть, введение аминазина ослабляет, по-видимому, чувство голода, т. е. блокада ретикулярной формации аминазином сказывается даже в субъективном исчезновении чувства голода.
В конце периода голодания десинхронизация в коре сохраняется, а в некоторых случаях несколько ослабевает, появляются небольшие группы альфа-активности в теменно-затылочных отделах коры. Быстрая острая активность в лобных отделах коры чаще всего сохраняется. Эти изменения могут быть связаны или с наличием «голодной» импульсации, которая к концу голодания снова усиливается, или является следствием сдвигов гуморальной среды — результатом прямого раздражения мозга «голодной» кровью.
Первый прием пищи приводит к появлению в ЭЭГ синхронных ритмов и исчезновению быстрой активности в передних отделах коры.
Когда больной начинает нормально питаться, в ЭЭГ регистрируется или синхронизированный фон, или десинхронизация сохраняется более длительное время, но отмечается некоторая тенденция к синхронизации.
Голодание вызывает возбуждение субкортикальных образований головного мозга. Это проявляется в развитии общей десинхронизации и появлении быстрой, острой активности в передних отделах коры, которая снимается аминазином и первым приемом пищи. Иногда в результате возбуждения глубоких структур мозга усиливаются и выявляются синхронные пароксизмальные разряды медленных и острых волн.
Отмеченные особенности изменения ЭЭГ выявляются у больных, у которых лечебное голодание и последующее питание в восстановительном периоде было связано со значительным терапевтическим эффектом.
Для примера приведем результаты исследования нескольких больных.
Больной П., история болезни № 2026, диагноз: циклотимия, депрессивное состояние. В больницу госпитализировался но собственному желанию с установкой на лечебное голодание. Проведено в течение 17 дней лечебное голодание с последующей молочно-растительной диетой, курс пироге-нала (3 инъекции внутримышечно от 2,5 до 7,5 гамм). Голодание перенес хорошо, но все время хотел есть. Уже в процессе голодания стало выравниваться настроение, стал активнее; в период питания наладился ночной сон. Появилось ощущение физического здоровья, почувствовал себя работоспособным. Выписан в хорошем состоянии. В течение одного года чувствовал себя здоровым, были нерезкие колебания настроения, с которыми справлялся самостоятельно.
Данные исследования ЭЭГ этого больного в динамике приведены на рис. 1.
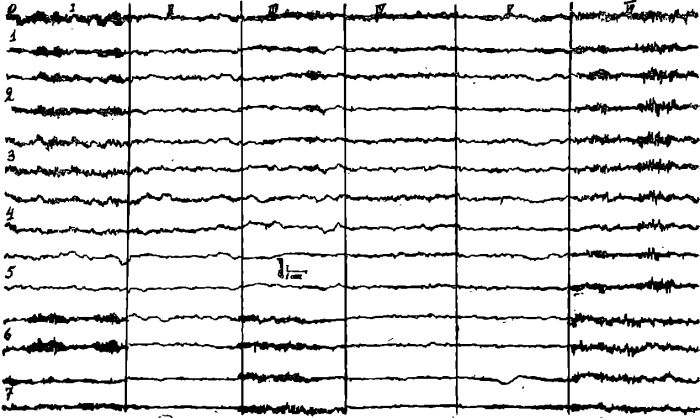
Рис. 1. ЭЭГ больного П.
Отведения:
1 — затылочное
2 — теменное
3 — центральное
4 — лобное
5 — височное
6 — теменно-лобное
7 — теменно-затылочное
I — фоновая запись; II — на 4 день голодания; III — через 20 минут после введения аминазина; IV — на 17 день голодания; V — через 15 минут после первого приема пищи; VI — на 10 день восстановительного периода.
До начала голодания в ЭЭГ выражен альфа-ритм, более регулярный с амплитудой до 70 мкв в затылочных и теменных отведениях, при биполярной записи амплитуда альфа-волн в теменно-лобных отведениях выше, чем в теменно-затылочных. В центральных и лобных отведениях небольшие группы альфа-волн сменяются бета-ритмом и низкоамплитудными синхронными группами тэта-волн. Бета-ритм и диффузные пологие медленные волны выявляются в височных отведениях. На 4 день голодания во всех отведениях выявляются десинхронизированные кривые, бета-активность наслаивается на диффузные пологие медленные волны; в лобных отведениях доминирует быстрая острая активность амплитудой 30—40 мкв. Через 20 минут после введения аминазина (25 мг внутримышечно) во всех отведениях появляются группы альфа-волн, быстрая активность в лобных отведениях исчезает. На 17 день голодания кривые остаются десинхронизированными, во всех отведениях доминирует бета-активность и быстрые острые волны амплитудой 30—40 мкв. Через 15 минут после первого приема пищи (стакан апельсинового сока) во всех отведениях отмечается замедление ритмов, исчезает быстрая острая активность, появляются небольшие группы низкоаплитудных альфа-волн. На 10 день питания, наряду с общей синхронизацией ЭЭГ, появлением групп заостренного альфа-подобного ритма во всех отведениях, сменяющегося группами бета-ритма, выявляются пароксизмальные синхронные разряды замедленных альфа и тета-волн амплитудой до 80—90 мкв, особенно четко выраженные в теменных, центральных лобных и височных отведениях.
Больной М. история болезни № 1419. Диагноз: шизофрения, депрессивно-параноидная форма, ипохондрический вариант. В больницу поступил с установкой на лечебное голодание. В течение 31 дня находился на режиме полного голодания с последующим переводом на растительно-молочный ахлоридный стол. На 17 день голодания значительно улучшилось самочувствие, уменьшились неприятные ощущения в теле. Состояние стало значительно лучше на 14—15 день питания. В течение одного года больной работал, чувствовал себя хорошо.
На ЭЭГ у этого больного (рис. 2) до лечения выявляется альфа-ритм преимущественно в затылочных и теменных отведениях, прерывающийся заостренными волнами.
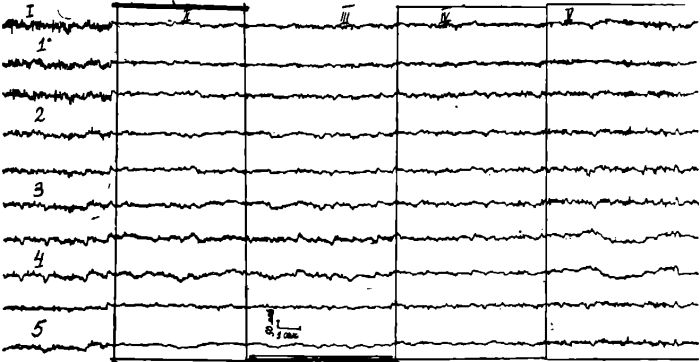
Рис. 2. ЭЭГ больного М.
Отведения те же, что и на рис. 1.
I — фоновая запись; II — на 5 день голодания; III — на 31 день голодания; IV— через 15 минут после первого приема пищи; V — на 14 лень восстановительного периода.
В остальных отведениях в фоне доминирует бета-ритм, сменяющийся группами альфа-подобных низкоамплитудных колебаний и пологих медленных волн. На 5 день голодания на ЭЭГ отмечается развитие десинхронизации, кривые во всех отведениях уплощены и в них бета-ритм наслаивается на пологие медленные волны различной частоты. В лобных отведениях выявляется быстрая острая активность небольшой амплитуды; На 31 день голодания ЭЭГ осталась без изменений, кривые во всех отведениях десинхронизированы, в лобных отведениях доминируют острые волны. Через 15 минут после первого приема пищи отмечается появление низкоамплитудных групп альфа-волн во всех отведениях и несколько усиливается бета-активность. Быстрая острая активность в лобных отведениях значительно ослабевает. На 14 день питания в ЭЭГ регистрируются небольшие группы альфа-воли, сменяющиеся группами бета-колебаний и синхронными вспышками острых волн, и тэта-волн преимущественно в темен-но-центральных отведениях.
Надо думать, что отмеченные изменения в ЭЭГ связаны с развитием при голодании возбудительных процессов в субкортикальных образованиях головного мозга и, в частности, в гипоталамических ядрах и в стволовой части ретикулярной формации. С этим может быть связана как общая десинхронизация, так и появление быстрой острой активности в передних отделах коры, более тесно связанными с неспецифическими структурами мозга. Прием пищи снимает возбуждение в подкорковых структурах, что и выражается в развитии синхронизации в ЭЭГ. Это было отмечено и в экспериментах на голодных кошках (8, 9, 10). По-видимому, при непродолжительном голодании затрагиваются не только холинергические механизмы головного мозга, но и адренэргические структуры. В наших наблюдениях введение на 3—4 день голодания аминазина в дозах 25 мг на кг веса приводило к появлению в ЭЭГ синхронизации, сменяющей исходную («голодную») десинхронизацию.
Здесь также интересно отметить, что регистрируемые изменения в ЭЭГ совпадают с фазами голодания и восстановительного периода (7) и соответствуют стадиям общего адаптационного синдрома (21): первая стадия — повышенной пищевой возбудимости (12—7 дней) — первая фаза общего адаптационного синдрома; вторая стадия — нарастание ацидоза и угнетение пищевой возбудимости, заканчивается ацидотическим кризом (7—13 до 17 дней) — вторая прешоковая фаза адаптационного синдрома; третья стадия компенсации или выравнивания (13—30 дней) — третья фаза равновесия общего адаптационного синдрома. Восстановительный период также имеет 3 стадии: первая стадия — нарастания пищевой возбудимости (4—5 дней), вторая стадия — интенсивного восстановления (2—3 недели), третья стадия — нормализации.
Иной характер носили изменения ЭЭГ у больных, у которых длительное голодание не дало значительного терапевтического эффекта.
По фоновой ЭЭГ этих больных можно разбить на две группы.
У первой группы больных (5 человек) и до начала голодания в ЭЭГ отмечалась десинхронизация. Эта десинхронизация может быть связана с наличием возбуждения в субкортикальных (возможно стволовых) образованиях головного мозга. И голодание может здесь вызвать перевозбуждение этих структур. Значительных изменений ЭЭГ в динамике голодания здесь не отмечается. И в начале, и в конце голодания, и после него ЭЭГ остается десинхронизированной. Введение аминазина в первые дни голодания вызывает появление синхронных групп альфа-волн.
Пример. Больная М., история болезни № 883. Диагноз: шизофрения, вяло текущая форма, неврозоподобный этап. В больницу поступила с установкой на лечебное голодание. Был проведен курс лечебного голодания в течение 28 дней, затем курс инсулина (32 комы), электросудорожная терапия — 4 сеанса, курс френалона, либриум. Терапевтического эффекта не отмечено.
На ЭЭГ исследуемой больной (рис. 3) до лечения отмечается десинхронизированный фон.
Во всех отведениях выявляется бета-ритм и диффузные низкоамплитудные острые волны, наслаивающиеся на пологие медленные колебания. На 4 день голодания кривые остались без изменения, но через 20 минут после введения аминазина (50 мг) появились синхронные группы альфа-колебаний, лучше выраженные в затылочных и теменных отведениях, и синхронные группы заостренных волн. В конце курса лечебного голодания кривые по существу остались без изменений, десинхронизированы, но несколько увеличилось количество диффузных пологих медленных волн. Такие же кривые сохранились и на 10 день восстановительного периода.
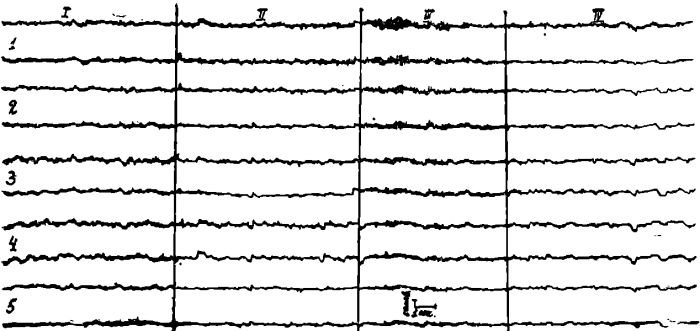
Рис. 3. ЭЭГ больной М. Отведения те же, что и на рис. 1.
I — фоновая запись; II — на 4 день голодания; III — через 20 минут после введения аминазина; IV — на 10 день восстановительного периода.
У второй группы больных с безрезультатным лечением голодом (4 человека) отмечается, напротив, не десинхронизированные, а гиперсинхронизированные фоновые кривые. В результате голодания изменений в ЭЭГ не выявляется. Десинхронизация не развивается и несколько усиливается гиперсинхронизация, увеличивается количество пароксизмальных разрядов высокоамплитудных медленных и острых волн, более четко выраженных в передних отделах коры.
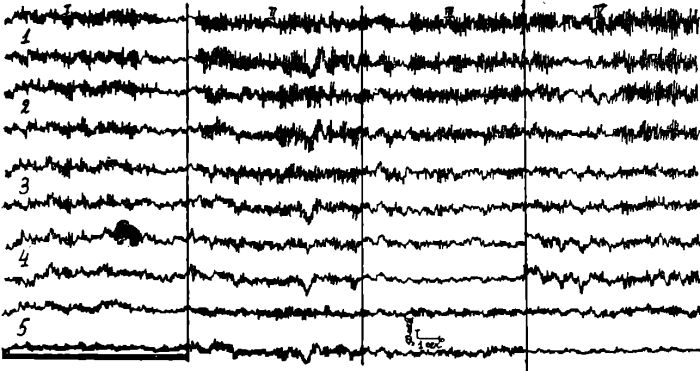
Рис. 4. Отведения и обозначения те же, что и на рис. 3.
Не исключено, что подобный фон ЭЭГ возникает в результате возбуждения неспецифических таламических ядер, которые, как считают некоторые авторы (20), находятся в антогонистических отношениях с ретикулярной формацией ствола мозга. В этом случае возбуждение ретикулярной формации при голодании не создается в результате этих антагонистических отношений.
Пример. Больной Г., история болезни № 1383. Диагноз: шизофрения, параноидная форма. В клинику поступил с установкой на лечебное голодание с целью «похудеть» и, как потом выяснилось, снять диагноз шизофрении. Больной в течение 28 дней воздерживался от пищи. «Голодал» по ареактивному типу без выраженных стадии. Выписан в прежнем состоянии.
На ЭЭГ у этого больного (рис. 4) до начала голодания отмечается гиперсинхронизированный фон за счет высокоамплитудного альфа-ритма, выраженного во всех отведениях.
Регионарные особенности ЭЭГ отсутствуют. Регистрируются высокоамплитудные синхронные разряды острых, а иногда и медленных волн. На 4 день голодания гиперсинхронизированный заостроенный альфа-ритм несколько усилился. После введения аминазина (25 мг внутримышечно) кривые остались без изменения. В конце голодания ЭЭГ не изменилась, осталась гиперсинхронизированной. С возобновлением питания и нормализацией его кривые ЭЭГ остались без изменений — гиперсинхронизированный альфа-ритм доминирует во всех отведениях.
Учитывая вышеизложенные факты можно сделать вывод, что в электроэнцефалограмме людей, находящихся на длительном лечебном голодании, развивается десинхронизация, появляются быстрые острые волны в лобных отделах коры и синхронные пароксизмальные разряды медленных и острых волн в передних отделах коры больших полушарий головного мозга. Указанные изменения ЭЭГ могут быть связаны с усилением восходящих активирующих влияний на кору большого мозга из субкортикальных образований, возможно из пищевых центров гипоталамических ядер и неспецифических структур мозгового ствола. Наряду с усилением восходящих активирующих влияний из холинергических структур гипоталамических ядер, которое отмечали некоторые авторы при кратковременном голодании у животных, уже в первые дни лечебного голодания выявляются восходящие активирующие влияния на кору из неспецифических структур ствола головного мозга. Во всяком случае, развившаяся в результате голодания десинхронизация в ЭЭГ человека и появившаяся быстрая острая активность в передних отделах коры снимается в первые дни голодания малыми дозами аминазина, который, как известно, блокирует не холинергический, а адренэргический субстрат ретикулярной формации. Надо отметить, что эти восходящие активирующие влияния на кору из возбужденных субкортикальных образований головного мозга подавляются при первом приеме пищи и постепенно исчезают при нормализации питания, когда исчезает возбуждение этих субкортикальных структур.
В порядке очень осторожного предположения можно думать, что благоприятный прогноз в результате проведения лечебного голодания следует ожидать в том случае, если в ЭЭГ больного развивается десинхронизация и появляется быстрая острая активность в передних отделах коры. Если же ЭЭГ больного и до начала голодания десинхронизирована или, напротив, гиперсинхронизирована, и в первые дни голодания десинхронизация не развивается, т. е. когда у больного имеется возбужденный фон ЭЭГ, очевидно, в результате уже имеющего место возбуждения, распространяющегося из глубоких образований головного мозга, то терапевтический аффект при лечебном голодании, возможно, будет неудовлетворительным.
ЛИТЕРАТУРА
1. Айрапетьянц Э. Ш. Высшая нервная деятельность и рецепторы внутренних органов, Л., 1952.
2. Айрапетьянц Э. Ш. В кн. Матер, научн. конф. по пробл. функциоиальн. взаимоотношения между различными системами организма в норме и патологии. Иваново, 1962, 503.
3. А и ох и н П. К- Ж. Высш. нервн. деятельн. им. Павлова, 1962, 12, 3, 379.
4. Д е л о в В. Е., Адамович Н. А., Б о р г е с т Г. П. Физиолог. ж. СССР им. Сеченова. 1961, 47, 9, 1083.
5. Д ы к м а и Л. М. В кн. Матер, иаучн. конференц. по пробл. физиолог. и патолог, кортико-висцеральн. взаимоотиошен. и функц. сист. организма. Иваново, 1965, 2, 598.
6. М а к а р о в П. О. Физиолог, ж. СССР им. Сеченова, 1952, 38, 3, 281.
7. Николаев Ю. С. Лечение шизофрении дозированным голоданием. Тр. ин-та психиатрии МЗ РСФСР, М., 1963, 09, 7.
8. Судаков К. В. Физиолог, ж. СССР им. Сеченова, 1962, 48, 2, 150.
9. Судаков К- В. Физиолог, ж. СССР им. Сеченова, 1965, 51, 4, 449.
10. Судаков К. В., Туренко А. И. В кн. Матер, научн. конференц. по пробл. физиолог, и патолог, кортико-висцеральн. взаимоотнош. и функц сист. организма. Иваново, 1965, 2, 321.
11. Толмасская Э. С. Бюлл. эксперимент, биолог, и медицин, 1948, 25, 1, 15.
12. Т о л м а с с к а я Э. С. О нервных механизмах координации соматических и висцеральных функций организма. М., 1964.
13. Т о л м а с с к а я Э. С., Д ы к м а н Л. М. Ж- высш. нервн. дея-тельи. им. Павлова, 1962, 12, 1, 161.
14. Ч е р н и г о в с к и й В. Н. Интероцепторы. М., I960.
15. Adam G., Meszoros J. C/R. Soc. Biol., 1958, 152, 1, 73.
16. Amass i an V. Fed. Pros., 1952, 11, 5.
17. A m a s s i а п V. J. Neurophysiol., 1951, 14, 435.
18. Dell P. J. Physiol., 1952, 44, 471.
19. F u 11 о n G. Physiology of the nervous system. N/.G. — Toronto, 1943.
20. Джаспер Г. В кн. Ретикулярная формация мозга. М., 1962, 237.
21. Селье Г. Очерки об адаптационном синдроме, М„ 1960.